نگاه کلی
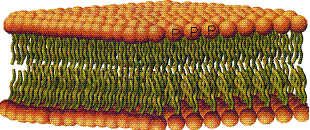
با همه مرزهای تفکیک شدهای که در سلول وجود دارد، یک قالب یا بسته سیتوپلاسمی تمام فضاهای موجود بین اندامکهایی را که بوسیله غشای سلولی احاطه شدهاند، پر میکند. این سیتوپلاسم زمینهای سیتوزول است. اگر با اولترا سانتریفوگاسیون مرحلهای تمام اندامکها حتی میکروزومها و ریبوزومها را هم از سیتوپلاسم جدا کنیم، بخشی شناور باقی میماند که همان سیتوزول است. سیتوزول بویژه در سلولهای در حال تمایز اهمیت خاصی دارد. |
ارگاستوپلاسم
در گذشته سیتوزول به صورت مادهای همگن در نظر گرفته میشد تا اینکه در اواخر قرن نوزدهم مشخص گردید که در برخی سلولها بویژه سلولهای ترشحی و سلولهایی که سنتز پروتئینی فعالی دارند، در بعضی قسمتهای سیتوزول باز دوستتر است و رنگهای بازی از جمله پیرونین را بهتر میپذیرد. به همین دلیل بخشهای باز دوست سیتوزول را سیتوپلاسم رنگ پذیر (کرومیدیال) مینامند. در سال 1887 گارنیر کلمه ارگاستوپلاسم را برای بخشهای بازوفیل سیتوپلاسم که به نظر او در بیوسنتز مواد نقش فعالی داشتند، بکار برد.ارگاستوپلاسم ، بخشهای باز دوستی نظیر ذرات نیسل موجود در جسم سلولی سلولهای عصبی ، سیتوپلاسم فعال و باز دوست سلولهای مخاطی و سلولهای ترشحی لوزوالمعده ، غدد بناگوشی ، سلولهای اصلی غدد معده و بخشهای باز دوست سلولهای کبدی را نیز شامل میشود. کاسپرین ، براشه و پژوهشگران دیگر نشان دادهاند که باز دوستی زیاد ارگاستوپلاسم به دلیل وجوداسیدهای ریبونوکلئیک است و به همین دلیل با تاثیر ریبونوکلئازها این باز دوستی از بین میرود.
از آنجا که اسیدهای ریبونوکلوئیک سیتوپلاسمی بویژه در ریبوزومها متراکمند، میتوان باز دوستی و فعال بودن سنتز پروتئینها در ارگاستوپلاسم را نتیجه فراوانی ریبوزومها در این بخش از سیتوزول دانست. در گذشته به جای سیتوزول بیشتر از کلمه هیالوپلاسم استفاده میشد که خود نشانهای از تصور همگن و شفاف بودن سیتوپلاسم زمینهای بوده است، تصوری که امروزه دگرگون شده است.
ترکیب تشکیل دهنده سیتوزول
درسیتوزول 85 درصد آب و حدود 15 درصد مواد مختلف موجود است. از این مواد بخش عمدهای را پروتئینها بویژه پروتئینهای آنزیمی اسیدهای آمینه ، گلوکز ، یونها ، mRNA ها ، tRNA ها و بطور خلاصه تمام مولکولهای لازم برای ایجاد انرژی و مواد لازم برای اعمال مختلف سلولی را شامل میشوند. پروتئینهای سازنده اسکلت سلولی از جمله توبولینها ، آکتینها ، میوزین ، تروپومیوزین و تروپونین نیز بخشی از پروتئینهای موجود در سیتوزول هستنند.برخی مواد موجود در سیتوزول میتوانند به نحوی تجمع یابند که به ساختمانهای قابل رویت با میکروسکوپ الکترونی تغییر شکل دهند. از جمله این ذرات ،گلیکوژن ، گویچههای لیپیدی و پروتئینهای اسکلت سلولی هستند که به صورت ریز لولهها و ریز رشتهها سازمان مییابند. یادآوری این نکته جالب است کهسانتریولها ، رشتههای دوک تقسیم و حتی تاژکها و مژکها زیر بنای ساختمانی ریز لولهای دارند.
 |
پروتئینهای موجود در سیتوزول
در سیتوزول پروتئینهای محلول ، پروتئینهای آنزیمی ، تمام آنزیمهای گلیکولیز ، آنزیمهای فعال کننده اسیدهای آمینه برای ورود به سنتز پروتئینها ، تمام ماشین سنتز پروتئینها و دیگر بخشهای محلول سیتوپلاسم وجود دارد. بطور کلی حدود 20 تا 25 درصد از کل پروتئینهای سلولی از جمله پروتئینهای آنزیمی در سیتوزول موجودند.آنزیمهای بسیاری از واکنشهای سلولی که به ATP نیاز دارند، TRNAها بخشهای دیگری از سیتوزول هستند. تغییر حالت سل به (ژل و به تبع آن تغییرات مختلفی از جمله تغییرات غلظتجنبش درون سلولی یا سیکلوز ، حرکت آمیبی ، تشکیل دوک تقسیم ، جابجایی کروموزومها ، برخی از تغییر شکلهای سلولی ، تسهیم یا شکافتگی سلول و مانند آن به سیتوزول وابسته است.
اندامکهای سیتوزولی
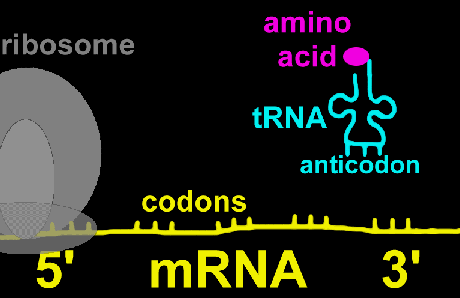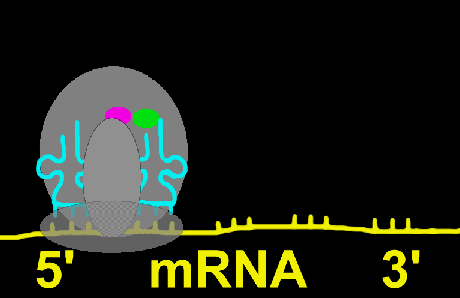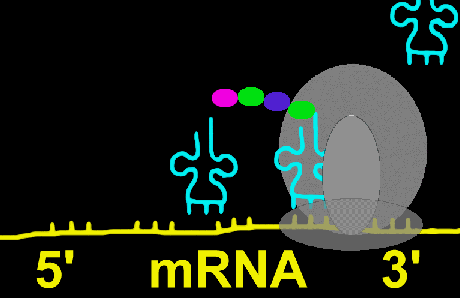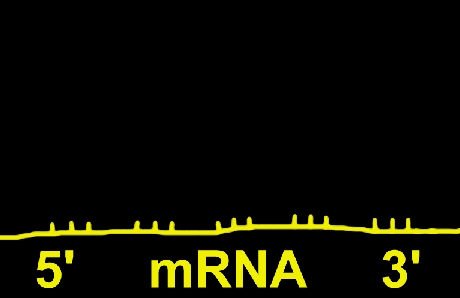
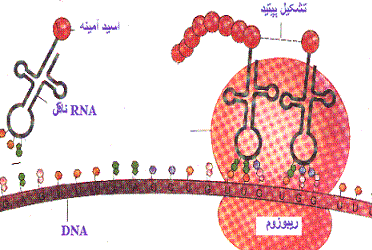
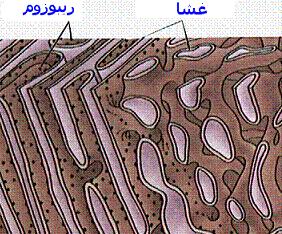
ریبوزوم
ریبوزومها دارای ترکیبات ریبونوکلئوپروتئین هستند و جایگاه کاتالیتیک برای سنتز پروتئین محسوب میشوند. ریبوزومها به صورت آزاد در سیتوزول و یا متصل به شبکه آندوپلاسمی دیده میشوند. ریبوزومها در سلولهای یوکاریوتی و پروکاریوتی دارای اندازه متفاوت هستند. ریبوزومها معمولا به صورت پلیزوم دیده میشوند که متشکل از تعداد زیادی ریبوزوم که توسط مولکولهای RNA پیک بهم متصل شدهاند و آماده پروتئین سازی هستند.شبکه آندوپلاسمی
شبکه آندوپلاسمی متشکل از لولههای درهم پیچیدهای است که ممکن است حاوی ریبوزوم بوده و به این علت شبکه آندوپلاسمی خشن یا دانهدار خوانده میشود و یا فاقد ریبوزوم باشد که در این صورت شبکه آندوپلاسمی صاف نامیده میشود. شبکه آندوپلاسمی محل اصلی سنتز پروتئین و چربی است و در تغییرات پس از سنتز این مولکولها نقش دارد. همچنین مولکولهای سمی را به مشتقات غیر سمی آنها که میتوانند از سلول خارج شوند تبدیل مینماید.دستگاه گلژی
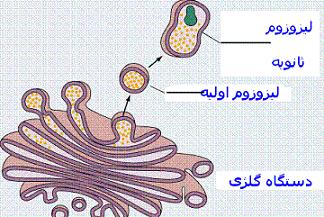
دستگاه گلژی به صورت اندامکهای شبکه مانند در نزدیکی هسته سلول قرار دارند. دستگاه گلژی از یک طرف با شبکه آندوپلاسمی و از طرف دیگر با وزیکولهای و لیزوزومها در ارتباط میباشد. عمل اصلی دستگاه گلژی پردازش و آماده سازی محصولات تازه سنتز شده سلولی و هدایت آنها به سوی سرنوشت نهاییشان میباشد.لیزوزوم
سادهترین ساختمانهای لیزوزومی لیزوزومهای اولیه هستند که عبارتند از کیسههایی با یک ماتریکس متراکم که اعتقاد بر این است که از گلژی مشتق شدهاند و حاوی آنزیمهای هیدرولیتیک هستند که جهت از بین بردن و هیدرولیز موادی که در گوارش سلولی از بین نرفتهاند بکار میروند. |



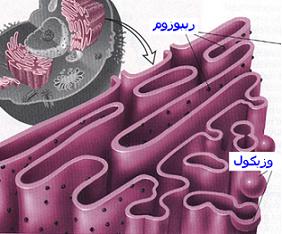



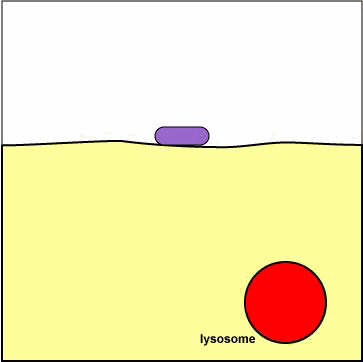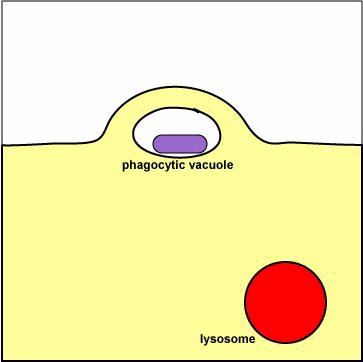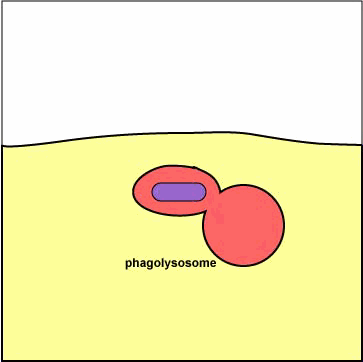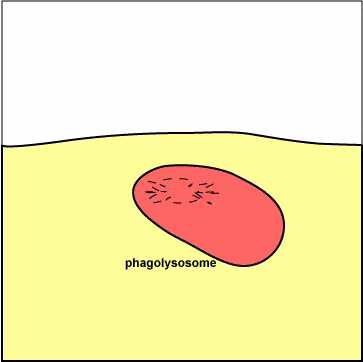
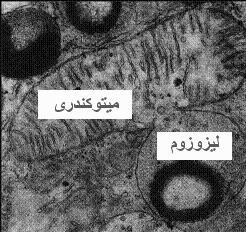
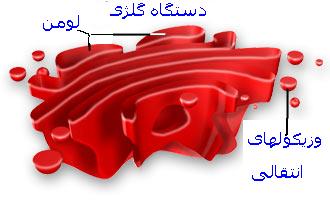 دستگاه گلژی
دستگاه گلژی