نگاه کلی
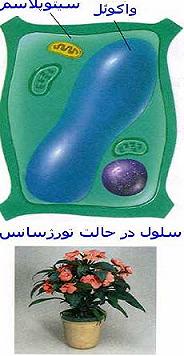
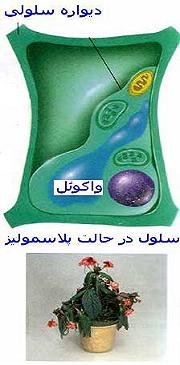
بررسی انواع مختلفی از بافتها نشان میدهد که بخشی از سیتوپلاسم بویژه در یاختههای گیاهی بوسیله اندامک حجیمی که آن را واکوئل مینامند پر شده است. مجموعه واکوئلهای هر یاخته ، دستگاه واکوئلی را تشکیل میدهد که آن را در مقایسه با کوندریوزومها (مجموع میتوکندریها) و پلاستیدوم (مجموع پلاستها) واکوئم مینامند. ممکن است واکوئلها 80 تا 90 درصد حجم یاختهای را پر کنند و سیتوپلاسم را به صورت لایه نازکی در کنارههای یاخته باقی گذارند.اولین گزارش در مورد واکوئلها بیشتر بر روی ویژگی شفاف بودن این اندامکها تکیه داشت و نام واکوئل از کلمه لاتین واکوئوس (فضای خالی) با این دید ابداع شد که واکوئل حفره یاختهای کم و بیش غیر فعال است. در سالهای اخیر ، پویایی و اهمیت تبادلهای واکوئلی به اثبات رسیده و واکوئلها به عنوان یکی از اندامکهای فعال یاختهای منظور شدهاند.
 |
تفکیک یا جدا سازی واکوئلی
عده زیادی از پژوهشگران واکوئلها را به صورت حفرههای آبکی که از تورم بخشهای کلوئیدی سیتوپلاسم بوجود آمدهاند، در نظر میگیرند. برخی دیگر آنها را نتیجه آبکی شدن محتوای بخشهایی از شبکه آندوپلاسمی دانستهاند. پس از پژوهشهای دووری مشخص شد که واکوئلها تشکیلات ساده موقتی نیستند، بلکه از بخشهایی مستقل و پایدار یاختهای هستند. وی با پلاسمولیز یاختهها در شرایط کم و بیش نامناسب موفق به تخریب سیتوپلاسم و حفظ واکوئلها شد.این تجربه را تفکیک یا جداسازی واکوئلی مینامند که موجب بدست آمدن حفرههایی شد که برای چند روز ویژگیهایی چون قدرت نگهداری رنگدانهها و توان تغییر حجم باز گشت پذیر با تغییر شرایط محیط خارجی را حفظ میکردند. این ویژگیها موجب این پندار شد که شیره واکوئلی بوسیله پوششی چسبنده ، ممتد ، قابل کشش ، قابل ارتجاع ، پایدار و دارای تراوایی نسبی احاطه شده است که دووری آن را تونوپلاست نام گذاشت. تمام این نتایج پس از کاربردمیکروسکوپ الکترونی ثابت گردیدند.
تغییرات واکوئلها
واکوئلها اندامکهایی دارای قابلیت تغییر و تحول هستند. تعداد ، اندازه ، نوع و غلظت محتوای درونی آنها بر حسب درجه تمایز یاختهای ، شرایط محیطی ، فصل و شرایط فیزیولوژیکی یاختهها تغییر میکند. با افزایش میزان تمایز یاختههای گیاهی ، واکوئلهای کوچک به تدریج بهم پیوسته و گسترش مییابند و واکوئل حجیمی را میسازند که بخش عمده یاخته را پر میکند و هسته و سیتوپلاسم را به کنارههای یاخته میراند.هنگام تمایز زدایی ، واکوئل حجیم چند بخش میشود. حجم این واکوئلها کاهش مییابد و موجب بازگشت سیتوپلاسم و هسته به وضعی مشابه یاخته جوان میگردد. واکوئلها اندامکهایی دارای تغییرات منظم نیز هستند. در یاختههای محافظ روزنه ، تغییرات واکوئلها دارای نظم شبانه روزی است. هنگام روز به دنبال افزایش فشار اسمزی که موجب تغییر شکل و حجیم شدن یاختهها میشود، روزنهها گشاد میشوند و شب هنگام که فشارها و اندازه واکوئلها کاهش مییابد، روزنهها تنگ میشوند.
 |
جنبشهای شبانه و حالت خواب اندامهای گیاهی (بسته شدن گلها ، تا شدن برگها هنگام شب ، باز شدن صبحگاهی آنها و نظایر آن) نیز نتیجه تغییرات فشار اسمزی یاختههایی است که در محلهای حساس قرار دارند. در یاختههای کامبیومی ، واکوئلها دارای نظم سالانه هستند. در زمستان کوچک شده و در بهار دوباره حجیم میگردند.
ساختار و فرا ساختار واکوئلها
ساختار واکوئل دو بخش اصلی شامل غشا و محتوای واکوئلی قابل تشخیص است. بررسیهای انجام شده بامیکروسکوپهای الکترونی فرا ساختار غشای واکوئلی یا تونوپلاست را بطور کلی مشابه پلاسمالم و متشکل از دو لایهفسفولیپیدی و پروتئینها نشان داده است. با این تفاوت که بخشهای گلوسیدی (قندی) گلیکولیپیدها در غشای واکوئل به طرف درون واکوئل قرار دارند و بخشی از این ساختارها به عنوان گیرنده برخی مواد موجود در واکوئلها عمل میکنند.محتوای واکوئلی
دستگاه واکوئلی دارای ترکیبات بسیار زیاد است که شامل یونهای کانی ، قندهای ساده و اولیگوزیدها ، اسیدهای آمینه ، اسیدهای آلی و دیگر (مثل اسد مالیک در ریشه واکوئلی سیب ، اسید اسکوربیک در مرکبات) پلی پپتیدها و پروتئینها و گلیکو پروتئینها ، موسیلاژهای پلی ساکاریدی و هتروزیدهای متنوع است. در مورد یونهای کانی ، تمام فنون جدید ، ورود انتخابی آنها را تایید میکنند. مخمرها تجمع واکوئلی قابل ملاحظهای از Mg+2 و فسفات دارند. برعکس سیتوپلاسم آنها دارای یونهای +K و +Na است.لولههای شیرابهای نیز مقدار زیادی Mg+2 دارند. در حالی که +K به غلظت برابر در واکوئل و سیتوزول آنها وجود دارد. آنیونهای واکوئلی مثل -Cl ، اغلب یونهای یک ظرفیتی هستند. محتوای واکوئلی مخزنی از ترکیبات پیچیده است که جنس و غلظت آنها بر حسب گونه ، نوع یاختهای و حالت فیزیولوژیکی جاندار بسیار متغیر است. برخی مولکولها بطور پایدار در واکوئلها ثابت شدهاند و برخی دیگر با سیتوپلاسم جابجایی دارند.
 |
این جنبشها اغلب دارای نظم هستند و در شرایط طبیعی میتوانند نوسانهای روزانه یا سالانه داشته باشند. مدت ذخیره مواد در واکوئلها بر حسب نوع یاخته متفاوت است و در بافتهای ذخیرهای طولانی است. برخی مولکولها مانند آنتوسیانها ، رنگدانههای مختلف ، اینولین و غیره تنها در شیره واکوئلی وجود دارند و برخی دیگر مثلساکارز ، مالات ، اسیدهای آمینه هم در واکوئل و هم در سیتوزول یافت میشوند. بنابراین درجه انتخاب واکوئل متغیر است.
محتوای واکوئلها ممکن است از مواد حد واسط فعالیتهای پایه متابولیسم اولیه یاخته باشند که ضمن جنبشهای سیتوپلاسمی کنار گذاشته شدهاند و یا محصولی از مسیرهای بیوسنتزی بسیار ویژه (متابولیسم ثانویه) هستند. از مهمترین محصولات متابولیسم اولیه موجود در واکوئلها میتوان به اسیدهای کربوکسیلیک ، گلوسیدها ، اسیدهای آمینه و پروتئینها اشاره کرد. محصولات متابولیسم ثانویه که در شیره واکوئلی وجود دارند شامل کومارین ، سیانوژنها ، فلاونوئیها ، تاننها ، آلکالوئیدها و از جمله آلکالوئیدهامرفین ، تئین چای ، کافئین قهوه ، کدئین خشخاش اشاره کرد.

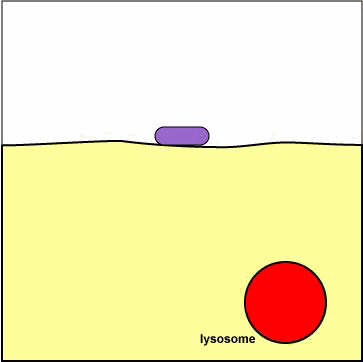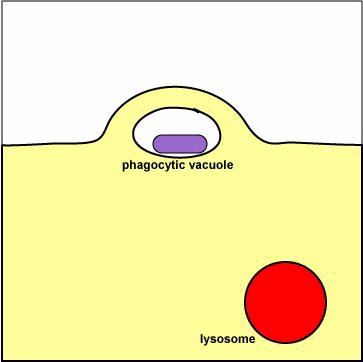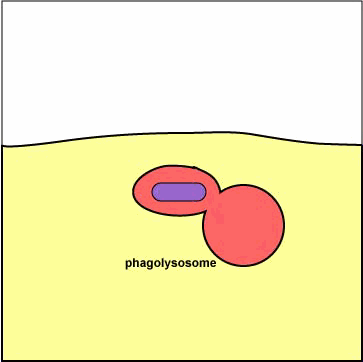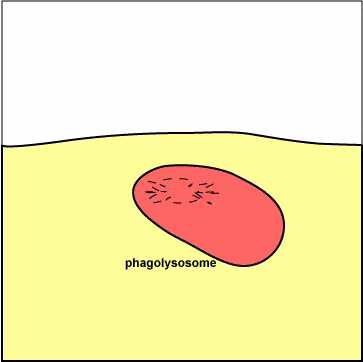
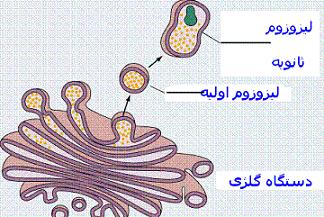
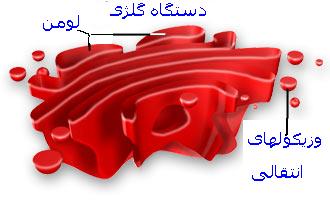 دستگاه گلژی
دستگاه گلژی