| مثلث برمودا محلی است وهمانگیز که در آن صدها هواپیما و کشتی در هوا و دریا ناپدید شدهاند. بیش از هزار نفر در این منطقه وحشت گم شدهاند، بدون اینکه حتی یک جسد یا قطعه پارهای از یک هواپیما یا کشتی مفقود شده ، به جا مانده باشد. |
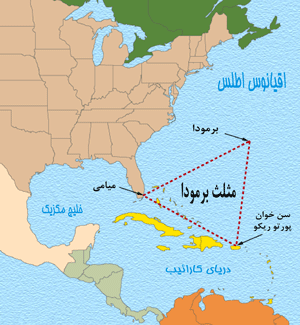 |
موقعیت مثلث برمودا
مثلث برمودا واقعا یک مثلث نیست، بلکه شباهت بیشتری به یک بیضی (و شاید هم دایرهای بزرگ) دارد که در روی بخشی از اقیانوس اطلس در سواحل جنوب شرقی آمریکا واقع است. راس آن نزدیک برمودا و قسمت انحنای آن از سمت پایین فلوریدا گسترش یافته و از پورتوریکو گذشته ، به طرف جنوب و شرق منحرف شده و از میان دریای سارگاسو عبور کرده و دوباره به طرف برمودا برگشته است. طول جغرافیایی در قسمت غرب مثلث برمودا 80 درجه است، بر روی خطی کهشمال حقیقی و شمال مغناطیسی بر یکدیگر منطبق میگردند. در این نقطه هیچ انحرافی در قطب نما محاسبه نمیشود.وینسنت گادیس که مثلث برمودا را نامگذاری کرده، آن را به صورت زیر توصیف میکند: « یک خط از فلوریدا تا برمودا ، دیگری از برمودا تا پورتویکو میگذرد و سومین خط از میان باهاما به فلوریدا بر میگردد. »
این محل فتنهانگیز و تقریبا باور نکردنی اسرار غیر قابل توصیف جهان را به خود اختصاص داده است. مثلث برمودا نامش را در نتیجه ناپدید شدن 6 هواپیمای نیروی دریایی همراه با تمام سرنشینان آنها در پنجم دسامبر 1945 کسب کرد. 5 فروند از این هواپیماها به دنبال اجرای ماموریتی عادی و آموزشی ، در منطقه مثلث ، پرواز میکردند که با ارسال پیامهایی عجیبی درخواست کمک کردند. هواپیمای ششم برای انجام عملیات نجات ، به هوا برخاست که هر شش هواپیما به طرز فوقالعاده مشکوکی مفقود شدند.
آخرین پیامهای مخابره شده آنها با برج مراقبت حاکی از وضعیت غیر عادی ، عدم روئیت خشکی ، از کار افتادن قطب نماها یا چرخش سریع عقربه آنها و اطمینان نداشتن از موقعیتشان بود. این در حالی بود که شرایط جوی برای پرواز مساعد بود و خلبانان و دیگر سرنشینان افرادی با تجربه و ورزیده بودند. با وجود مدتها جستجو هیچ اثری از قطعه شکسته ، لکه روغن ، آثاری از اجسام شناور ، خدمه یا تجمع مشکوکی از کوسهها دیده نشد. هیچ حادثهای چه قبل و چه بعد از آن ، تا این حد حیرتآورتر از ناپدید شدن دسته جمعی هواپیماهای مذکور نبوده است. در حوادثی مشابه در این منطقه قایقها و کشتیهایی مفقود شدهاند (قربانیان مثلث برمودا)، در برخی موارد هم فقط خدمه و سرنشینان ناپدید گشتهاند.
منطقه وحشت
همه روزه هواپیماهای متعددی بر فراز مثلث برمودا پرواز میکنند. کشتیهای بزرگ و کوچک در آبهای آن در حال تردند و افراد زیادی برای بازدید ، به این منطقه مسافرت میکنند، بدون آنکه اتفاقی بیفتد. از طرف دیگر ، در دریاها و اقیانوسها در سراسر دنیا ، کشتیها و هواپیماهای زیادی مفقود شده و میشوند. پس چرا فقط مثلث برمودا از بقیه مناطق تفکیک شده است. علت این است که اولا هیچ امیدی برای یافتن حتی اثر و نشانهای وجود ندارد. ثانیا در هیچ منطقه دیگر چنین ناپدید شدنهای بی دلیل ، بیشمار و نامعلوم روی نداده و به این خوبی ثبت نشده است.مشاهدات و گزارشات
در بیشتر اتفاقات مثلث برمودا ، اکثر هواپیماها در حالی ناپدید شدهاند که تماس رادیویی خود را با ایستگاههای مبدا و مقصدشان تا آخرین لحظه حفظ کردهاند و یا برخی دیگر در لحظات آخر پیامهای غیر عادی مخابره کردهاند که حاکی از عدم کنترل آنان بر روی دستگاه و ابزارها بوده است و یا چرخش عقربههای قطب نما به دور خود و تغییر رنگ آسمان اطراف به زردی و مه آلودی ، آن هم در روز صاف و آفتابی و یا تغییراتی غیر عادی در آبها که تا لحظاتی قبل آرام بودهاند، بدون بیان هیچ دلیل روشنی از چگونگی این وقایع.این پیامها رفته رفته ضعیفتر و غیرقابل تشخیصتر شده و یا سریعا قطع شدهاند. دقیقا مثل اینکه چیزی ارتباط رادیویی را قطع کرده باشد و یا چنانچه اظهار عقیده شده، در حال دور شدن و عقب رفتن از فضا و زمان بوده و دورتر و دورتر شدهاند. در برخی موارد گزارشها حاکی از آن بود که نوری ناشناخته و غیر قابل تشریح روئیت شده است. همچنین توده سیاه و تاریکی در سطح دریا که پس از مدتی ناپدید شده ، در جریان اتفاقات مزبور گزارش شده است.
در مواردی هم گزارش شده که نقطه تاریک بزرگی در میان ستارگان در آسمان دیده شده که نوری متحرک از طرف زمین به آن قسمت وارد شده و سپس هر دو ناپدید شدهاند. در تمام مدت دیده شدن تاریکی ، دستگاهها و سایر ابزارهای قایقهای ناظر از کار افتاده بودند که پس از رفع تاریکی آسمان ، دوباره شروع بکار کردهاند.
در یک مورد هم پیامی عجیب از یک کشتی باری ژاپنی بدین مضمون دریافت گردید. "خطری همانند یک خنجر هم اکنون ... به سرعت میآید ... ما نمیتوانیم فرار کنیم ..." در هر حال بدون اینکه مشخص شود خنجر چه بود، کشتی ناپدید شد.
علل واقعه
علل فرضی طبیعی
توضیحات و علل فرضی مختلفی درباره حوادث مثلث برمودا ارائه شده است که معمولترین فرضیات بر اساس مرگ غیر طبیعی (زیرا هیچ جسدی تا کنون بدست نیامده است.) بنا شده است. این توضیحات عبارتند از:جزر و مد ناگهانی دریا در نتیجه زلزله در اعماق دریا ، وزش بادهای مخرب و اختلالات جوی ، گویهای آتشفشان که موجب انفجار هواپیماها میشود، گرفتار آمدن در جاذبه یک گرداب یا گردباد که باعث سقوط و انهدام هواپیماها یا انحراف مسیر کشتیها و مفقود شدن آنها در آب میشود، تحت تاثیر نیرویی مغناطیسی قرار گرفتن و اختلالات امواج الکترومغناطیسی، ولی این دلایل توجیه قابل قبولی برای ناپدید شدن هواپیماها و کشتیهای متعدد در یک منطقه نیست.
علل فرضی غیر طبیعی
دستگیری و ربوده شدن به وسیله زیردریایی یا بشقاب پرندههایی متعلق به کراتی دیگر که برای تحقیق درباره حیات و زندگی باستان و حال ما انسانها به کره زمین آمدهاند، میتواند علتی غیر طبیعی برای توجیه وقایع باشد.یکی از عجیبترین پیشنهادات در این مورد بوسیله ادگار کایس ، پیشگو و روانکاو و حکیم در دهه پنجم قرن بیست ، ارائه شده است. به عقیده وی قرنها قبل از کشف اشعه لیزر ، بومیان سواحل اقیانوس اطلس از کریستال به عنوان یک منبع انرژی و قدرت استفاده میکردند. به نظر کاین نوعی نیروی شیطانی القا شده از سوی آنها ، در عمق یک مایلی در قسمت غرب اندروس غرق شده که هنوز در برخی مواقع باعث از کار انداختن ابزار و وسایل الکتریکی کشتیها ، هواپیماها و در نهایت نابودی آنها میگردد.
ام. ک. جساپ که یک فضانورد ، منجم و متخصص کره ماه است، در کتابش به نام « در مورد بشقاب پرندهها » ابزار میدارد که ناپدید شدن کشتیهای مشهور در مثلث برمودا ، به وسیله اجسام پرنده صورت گرفته است. وی مفقود شدن خدمه آنها را نیز به اجسام مزبور ربط میدهد. به عقیده جساپ یوفوها هر چه هستند، حوزه مغناطیسی موقتی ایجاد میکنند که دارای طرحی یونیزه شده است و میتواند باعث متلاشی شدن یا ناپدید شدن هواپیماها و کشتیها گردد. او روی این سوال کار میکرد که چگونه نیروی مغناطیسی کنترل شده و میتواند باعث نامرئی شدن گردد. نظریه میدان واحد انیشتین او را مجذوب کرده بود. جساپ هر دو اینها را کلیدی میدانست برای ظهور و محو شدن ناگهانی بشقاب پرندهها و ناپدید شدن کشتیها و هواپیماها. ولی مرگ امکان ادامه فعالیت و نتیجه گیری را از جساپ گرفت و تحقیقاتش نیمه تمام ماند.
داستانی عجیب
حادثهای در اثر اختلال زمانی در فرودگاه میامی رخ داد که هرگز توضیحی قابل قبول برای آن وجود نداشته است. این واقعه مربوط به یک هواپیمای مسافربری بود که برای فرود در باند آماده بود و با رادار مرکز کنترل هوایی ردیابی میگردید که ناگهان ده دقیقه از صفحه رادار ناپدید شد و سپس دوباره ظاهر گشت. هواپیما بدون هیچ واقعهای فرود آمد و خلبان و خدمه از آنچه افراد پایگاه میگفتند ابراز تعجب کردند، زیرا تا آنجا که به خدمه مربوط میشد، هیچ اتفاق غیر عادی نیفتاده بود. جالب این که ساعتهای همه آنها حدود ده دقیقه از زمان واقعی عقبتر بود. در حالی که هواپیما درست 20 دقیقه قبل از این واقعه وقت اصلی را کنترل کرده بود و در آن هنگام هیچ اختلاف زمانی وجود داشت.- آیا مثلث برمودا و نقاط مشخص دیگر به صورت ماشینی عظیم عمل میکنند تا اختلالاتی بوجود آورند؟
- آیا آنها میتوانند گردابهایی را چه در داخل و چه در خارج از جو بوجود آورند که اجسام و اشیا به داخل آنها بیفتد و به بعد زمان و مکانی دیگر منتقل شوند؟
گذشته و آینده برمودا
به نظر میرسد که این منطقه طی زمانهای متمادی گذشته نیز در افسانهها به منزله مکانی ترسناک وجود داشته و حتی خیلی قبل از تاریخ کشف آن و بعد از آن تاریخ تا صدها سال با عناوین «دریایی از مقبرهها» ، «مثلث شیطان» ، «مثلث مرگ» ، «دریای بدبختی» ، «گورستان آتلانتیک» نامیده میشده است.شومی و بدشگونی مثلث برمودا حتی در عصر فضا نیز باعث تعجب انسانهایی چون کریستف کلمب و فضانوردان آپولو 13 که یکی کاشف در زمین و دیگری در فضاست، شده است.
اینکه چرا وقایع عجیب این منطقه گزارش نمیشود، شاید به دلیل ایجاد رعب و وحشت عمومی باشد، شاید هم چون دلیل اصلی وقایع معلوم نیست، اتفاقات مربوطه بازتاب نمییابد. البته در اغلب گزارشات ارائه شده هم سانسورهایی وجود دارد که اصل وقایع را سرپوشیده نگه میدارد.
- آیا این مثلث دوباره قربانیان دیگری میگیرد؟
- آیا بشر موفق به کشف راز آن خواهد شد؟